Chúng ta đã biết hiện tượng cộng hưởng từ và hiệu ứng cộng hưởng từ của các hạt proton (hạt nhân nguyên tử hydro). Trong cơ thể sống, hydro có mặt trong thành phần của nhiều hợp chất thiết yếu, đặc biệt là nước và mỡ. Vì vậy, nghiên cứu tính chất cộng hưởng từ của proton có vai trò cực kỳ quan trọng trong quá trình tạo ảnh cộng hưởng từ và là bước đầu tiên giúp chúng ta hiểu được quá trình này.
Chúng ta biết rằng nước là một chất thiết yếu của mọi cơ thể sống và thường chiếm một hàm lượng cao trong tế bào. Ngoài việc tham gia vào cấu trúc của tế bào, nước còn là môi trường để các quá trình sống xảy ra và trong nhiều trường hợp còn tham gia trực tiếp vào các quá trình sống. Với một mật độ cao và một vai trò cực kỳ quan trọng như thế, nước là thành phần chính tạo ra các tín hiệu cộng hưởng từ. Ngoài nước, mỡ cũng là một nguồn tín hiệu cộng hưởng quan trọng do chứa các nguyên tử hydro ở những vị trí “thuận lợi”.
Trong phần này chúng ta sẽ tập trung tìm hiểu về hiệu ứng cộng hưởng từ của các mô cơ thể. Nội dung của phần bao gồm:
- Độ xê dịch hóa học
- Đặc tính cộng hưởng từ của nước
- Thời gian hồi giãn dọc T1
- Thời gian hồi giãn ngang T2
1. ĐỘ XÊ DỊCH HÓA HỌC
Như chúng ta đã biết, tần số cộng hưởng hay tần số quay f của proton phụ thuộc vào từ trường ngoài B0 và hằng số Larmor g qua phương trình Larmor:
f = γB0
Khi tham gia cấu thành các phân tử, tần số cộng hưởng của proton còn phụ thuộc vào liên kết hóa học giữa proton với các nguyên tử khác và vị trí của proton trong phân tử. Sự khác biệt giữa tần số cộng hưởng của proton do liên kết hóa học và vị trí của proton trong phân tử được gọi là độ xê dịch hóa học (chemical shift).
Thực tế, độ xê dịch hóa học biểu thị sự chênh lệch tương đối giữa tần số quay của các proton trong các phân tử khác nhau, được tính bằng đơn vị ppm (parts per million) hay đơn vị phần triệu. Chẳng hạn, độ xê dịch hóa học giữa nước và mỡ là 3,5 ppm, nghĩa là proton trong nước quay nhanh hơn proton trong mỡ khoảng 3,5 phần triệu.
Khi cường độ từ trường tăng lên, tần số quay của các proton cũng tăng theo. Proton có tần số quay càng nhanh sẽ bị ảnh hưởng nhiều hơn và quay càng nhanh hơn. Nghĩa là sự khác biệt tuyệt đối giữa tần số quay của proton ở hai loại phân tử khác nhau có thay đổi theo cường độ từ trường nhưng độ xê dịch hóa học của chúng không thay đổi.
Liên kết hóa học của proton
Chúng ta biết rằng phân tử nước H2O có hai nguyên tử hydro (proton) và một nguyên tử oxy. Nguyên tử oxy tham gia hai liên kết hóa học với các nguyên tử hydro, một liên kết cho mỗi hydro. Trong mỗi liên kết, nguyên tử oxy và hydro đều đóng góp một điện tử vào đám mây chung (Hình 1a).
Tương tự, trong các phân tử lipid với các gốc methylene CH2 và methyl CH3, nguyên tử carbon cũng tham gia liên kết hóa học với các nguyên tử hydro bằng cách đóng góp một điện tử vào đám mây chung giữa carbon và hydro. Ngoài ra, mỗi nguyên tử carbon còn có thể tham gia vào các liên kết hóa học với hai nguyên tử carbon khác như được minh họa trong Hình 1b.
Độ xê dịch hóa học giữa nước và mỡ
Khi so sánh proton trong phân tử nước H2O và proton trong gốc CH2 hoặc CH3 của lipid, chúng ta thấy rằng ái lực với điện tử của oxy cao hơn của car- bon: oxy của nước giữ đám mây điện tử chung của nó với hydro chặt hơn so với carbon của lipid.
Mặt khác, điện tử vốn mang điện âm, khi quay quanh trục của mình cũng sinh ra một từ trường. Khi điện tử ở gần proton, từ trường này tác động vào proton và làm thay đổi chút ít tần số quay của proton.
Với một từ trường cố định, proton của nước bị ảnh hưởng của điện tử ít hơn so với proton của lipid, và vì vậy proton của nước quay hơi nhanh hơn (Hình 1). Khi từ trường mạnh hơn, proton của nước càng quay nhanh hơn, chênh lệch thực sự về tần số quay của nước và mỡ sẽ lớn hơn.

Hình 1: Liên kết hóa học của proton trong phân tử nước và lipid. (a) Đám mây điện tử chung giữa oxy và hydro bị lệch nhiều về phía oxy, cho phép pro- ton quay nhanh hơn. (b) Đám mây điện tử giữa carbon và hydro trải đều giữa chúng làm cho proton trong các phân tử lipid quay chậm hơn.
Chúng ta cũng cần biết rằng do lipid là loại phân tử có cấu trúc phức tạp nên thực chất tần số quay của proton trong lipid cũng khác nhau tùy theo vị trí của chúng. Nghĩa là bản thân mỡ cũng có độ xê dịch hóa học nội tại của mình. Nói cách khác, mỡ là một chất có độ xê dịch hóa học không đồng nhất.
Ảnh hưởng của độ xê dịch hóa học
Như chúng ta đã biết, tín hiệu cộng hưởng từ phụ thuộc vào “sức mạnh tổng hợp” của các proton. Nghĩa là sau khi tắt xung kích thích, số proton quay cùng pha càng lớn, tín hiệu cộng hưởng từ càng mạnh.
Độ xê dịch hóa học, vốn là sự khác biệt về tần số quay của các proton do đặc thù hóa học của các chất, sẽ nhanh chóng làm cho các proton “xa rời nhau”. Hiện tượng suy giảm cảm ứng tự do khi đó xảy ra rất nhanh, đồng nghĩa với việc tín hiệu cộng hưởng từ nhanh chóng giảm cường độ và biến mất.
Độ xê dịch hóa học còn khiến chúng ta phải tạo ra một xung kích thích có tần số cộng hưởng thích hợp, và đủ bao quát để cộng hưởng được với những proton có tần số cộng hưởng khác nhau. Điều này lại tạo ra một từ trường kém đồng nhất và đến lượt nó, từ trường kém đồng nhất này lại làm mất nhanh tín hiệu cộng hưởng từ.
Tuy nhiên trong một số trường hợp, độ xê dịch hóa học được tận dụng tối đa nhằm làm cho vùng cần khảo sát dễ thấy hơn. Chẳng hạn bằng cách dùng một xung kích thích có tần số thích hợp, người ta có thể làm mất hẳn tín hiệu được phát ra từ các mô mỡ, chỉ còn lại tín hiệu được phát ra từ nước. Nói cách khác, chúng ta đã “xóa mất mỡ” của vùng cần khảo sát trên hình cộng hưởng từ.
Về sau qua từng bối cảnh cụ thể, chúng ta sẽ bàn luận chi tiết hơn về ảnh hưởng của độ xê dịch hóa học và giải thích rõ hơn ý nghĩa của từng điểm đã nêu ở trên.
2. ĐẶC TÍNH CỘNG HƯỞNG TỪ CỦA NƯỚC
Nước có mặt trong tất cả các mô cơ thể. Trong tế bào, nước là thành phần chủ yếu của bào tương, làm dung môi để hòa tan các chất và là môi trường để các quá trình sống xảy ra. Ngoài tế bào, nước có mặt trong các khoảng gian bào, mô kẽ và là thành phần chủ yếu của các chất dịch và máu.
Nhờ sự có mặt ở khắp nơi với mật độ cao và một cấu trúc hóa học với hai proton (hạt nhân hydro), tín hiệu cộng hưởng từ của nước từ các mô khác nhau trong cơ thể là nguồn tín hiệu quan trọng nhất để từ đó chúng ta xây dựng các ảnh cộng hưởng từ. Trong thực tế lâm sàng, các kỹ thuật phát xung và đo đạc tín hiệu chỉ cho phép kích thích và ghi nhận các tín hiệu được phát ra từ nước và mỡ. Do vậy đặc tính cộng hưởng từ của nước là một đề tài quan trọng cần được chú ý đặc biệt.
Mô hình hai khoang
Mặc dù có mặt trong hầu hết các mô với nhiều cấu trúc phức tạp khác nhau nhưng về mặt cộng hưởng từ, chúng ta có thể dùng một mô hình hai khoang đơn giản để mô tả tình trạng của nước: nước tù (bound water) và nước tự do (free water).
Nước tù nói chung là phần nước trong cơ thể bị hạn chế chuyển động. Tình trạng này xảy ra do các phân tử nước bị hút và bị giữ chặt hơn khi chúng nằm gần các phân tử cực lớn như protein, phospholipid và ADN. Những đại phân tử này thường có nhiều điểm tích điện trên bề mặt và có khuynh hướng hút các proton mang điện tích dương. Chúng tạo thành các cầu nối lỏng lẻo làm giảm bớt khả năng chuyển động tự do của các phân tử nước.
Nước tự do là phần nước trong cơ thể có thể chuyển động tự do vì không nằm gần các đại phân tử hoặc trong môi trường không có hoặc có ít các đại phân tử. Nếu môi trường có các đại phân tử, nước tự do bị chúng hút rất yếu nhờ nằm cách xa chúng và lực hút này dễ dàng bị phá vỡ do tốc độ di chuyển của chính các phân tử nước.
Nước trong các mô
Trong các mô mềm như gan, tụy và não với diện tích nội bào lớn, lượng nước tù chiếm nhiều hơn nước tự do. Ngược lại trong dịch não tủy, máu và nước tiểu, lượng nước tự do rõ ràng là chiếm đa phần. Nang và tuyến cũng có lượng nước tự do nhiều hơn đáng kể so với nước tù. Ở mức trung gian, các chất dịch với tỷ lệ protein cao như dịch tiết và dịch khớp cũng làm giảm đáng kể tỷ lệ lượng nước tự do trong dịch.
Đặc tính cộng hưởng từ
Trước tiên chúng ta cần nhấn mạnh rằng proton trong các phân tử nước tù và nước tự do đều có tần số cộng hưởng như nhau, nghĩa là không có độ xê dịch hóa học giữa nước tù và nước tự do. Khác biệt giữa chúng nằm ở chỗ: nước tự do chuyển động nhiều hơn nên tác động của từ trường đối với chúng xảy ra chậm hơn. Ngược lại, do bị các đại phân tử giữ chặt hơn, nước tù cùng với các đại phân tử hình thành một môi trường có cấu trúc khá “ổn định”. Sự ổn định này khiến cho nước tù “nhạy cảm” hơn với sự thay đổi của từ trường ngoài.
Chính vì vậy, tỷ lệ giữa nước tù và nước tự do trong các mô có ảnh hưởng rất lớn đến các thời gian hồi giãn đặc trưng của các mô. Hai phần tiếp theo sẽ bàn luận chi tiết về các đặc tính thời gian cùng với ảnh hưởng của tỷ lệ nước tự do và nước tù trong các mô đối với những đặc tính đó.
3. THỜI GIAN HỒI GIÃN DỌC T1
Khi không có từ trường ngoài tác động, các proton trong môi trường tự quay quanh trục của mình. Hướng của các trục quay hoàn toàn ngẫu nhiên nên mỗi proton mặc dù có một từ trường riêng nhưng tổng từ trường chung của các proton ở trạng thái này vẫn bằng zero.
Khi có một từ trường ngoài B0 thật mạnh tác động vào, các proton trong môi trường bắt đầu định lại trục quay của chính mình cho phù hợp với hướng tác động của từ trường. Theo quy ước, chúng ta xem như hướng tác động này là hướng của trục z. Trục quay của các proton có thể hướng cùng chiều với trục z hoặc ngược chiều với z. Số proton có trục hướng cùng chiều với z chỉ nhiều hơn chút ít so với số proton hướng ngược chiều trong mỗi triệu proton. Tuy khác biệt này rất nhỏ nhưng do số lượng proton trong cơ thể rất lớn (có ý nghĩa nhất là các proton nằm trong nước và mỡ), chúng vẫn tạo ra một từ trường nhỏ gọi là độ từ hóa thực M0 có hướng vectơ (hướng tác động) cùng chiều với chiều vectơ B0 (Hình 2).
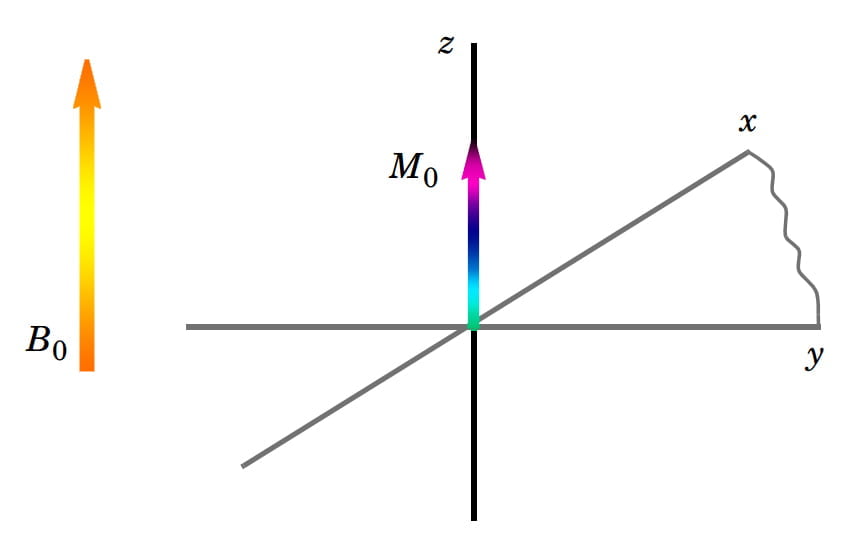
Hình 2: Độ từ hóa thực M0 hướng theo trục z được tạo thành do sự chênh lệch về số lượng giữa các proton có trục quay cùng và ngược chiều với B0.
Trong bối cảnh này, độ từ hóa thực cũng chính là độ từ hóa dọc.
Khi phát ra một xung kích thích RF quay quanh trục z có tần số quay bằng với tần số quay của proton, từ trường B1 do xung RF tạo ra sẽ kéo vectơ M0 lệch khỏi trục z một góc lật a. Khi a = 90o, vectơ M0 bị lật hoàn toàn vào mặt phẳng xy, tạo thành độ từ hóa ngang.
Lúc này nếu tắt xung kích thích, môi trường chỉ còn chịu tác động của từ trường B0. Bối cảnh này tương tự như bối cảnh khởi đầu khi áp đặt từ trường B0 khi các proton đang có trục quay ngẫu nhiên định lại trục quay để rồi dần dần tạo ra độ từ hóa thực M0 hay độ từ hóa dọc. Nghĩa là sau khi tắt xung, dưới tác động của B0, độ từ hóa dọc dần dần được hình thành trở lại. Quá trình này gọi là quá trình hồi giãn dọc (longitudinal relaxation).
Đường cong của quá trình hồi giãn dọc
Trong quá trình hồi giãn dọc, vectơ từ hóa dọc lớn dần, lúc đầu thật nhanh nhưng về sau chậm dần cho đến độ lớn tối đa của nó là M0. Nghĩa là quá trình hồi giãn dọc xảy ra từ từ, ban đầu nhanh rồi dần dần chậm lại. Để cho dễ hình dung, người ta thường mô tả quá trình hồi giãn dọc bằng một đường cong cho thấy mức độ hồi phục của vectơ từ hóa dọc theo thời gian (Hình 3). Đường cong này cho thấy rằng hơn phân nửa độ lớn của vectơ từ hóa dọc được hồi phục rất nhanh, và người ta chọn thời gian hồi phục được khoảng 63% độ lớn của M0 làm thời gian tiêu biểu cho quá trình hồi phục này và gọi nó là thời gian hồi giãn dọc hay thời gian T1.
Hình 3 cũng cho thấy rằng khôi phục hoàn toàn M0 cần phải mất một khoảng thời gian gấp bốn đến năm lần thời gian T1. Do vậy trong thực tế, T1 được dùng thay cho thời gian hồi giãn dọc thực sự. Nếu dùng đơn vị đo là ms (mili-giây), thời gian T1 có giá trị từ khoảng 100 ms đến 3000 ms (3 giây).
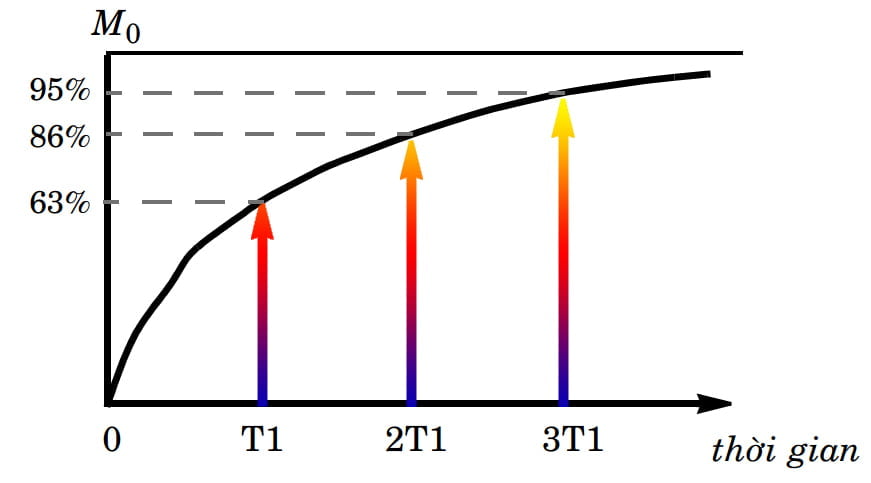
Hình 3: Đường cong mô tả thời gian hồi giãn dọc cho thấy mối liên hệ giữa độ lớn của vectơ từ hóa dọc với thời gian. T1 là thời gian khôi phục được khoảng 63% độ lớn so với độ từ hóa thực M0. Sau khi mất thêm một khoảng thời gian T1 nữa (2T1), vectơ từ hóa dọc khôi phục được khoảng 86%. Thực tế, để khôi phục hoàn toàn độ từ hóa dọc phải mất một khoảng thời gian gấp bốn hoặc năm (4T1 hoặc 5T1) lần T1.
Thời gian T1 của một số mô điển hình
Với một từ trường có cường độ nhất định, mỗi mô trong cơ thể đều có một thời gian T1 khá đặc trưng. Trong các mô khác với mô mỡ, thời gian này thường phụ thuộc vào tỷ lệ giữa nước tự do và nước tù: nước tự do càng nhiều, thời gian T1 càng dài vì sự chuyển động của các phân tử nước làm cho các proton khó định hướng lại trục quay của mình dưới tác dụng của từ trường ngoài. Chẳng hạn với từ trường 1,5 Tesla, thời gian T1 của dịch não tủy (nhiều nước tự do) có thể xấp xỉ 2400 ms, trong khi đó thời gian T1 của các mô mềm như chất trắng và chất xám thường không quá 1000 ms. Hình 4 trình bày một bảng các giá trị thời gian T1 ghi nhận được của các mô khác nhau trong một nghiên cứu được tiến hành và so sánh giữa các từ trường 0,5 và 1,5 Tesla.
Ghi nhận tổng quát từ Hình 4 cho thấy rằng mô mỡ có thời gian T1 ngắn hơn hẳn so với các mô khác. Các mô mềm như gan, cơ do diện tích bề mặt nội bào lớn và cấu trúc mô nhất quán nên chúng có thể giữ chặt được nhiều phân tử nước và làm cho thời gian T1 của chúng cũng khá ngắn. Dịch não tủy với thành phần chủ yếu là nước tự do có T1 rất dài. Khi so sánh giữa chất xám và chất trắng, T1 của chất trắng ngắn hơn so với T1 của chất xám vì chất trắng chứa nhiều mỡ hơn chất xám.
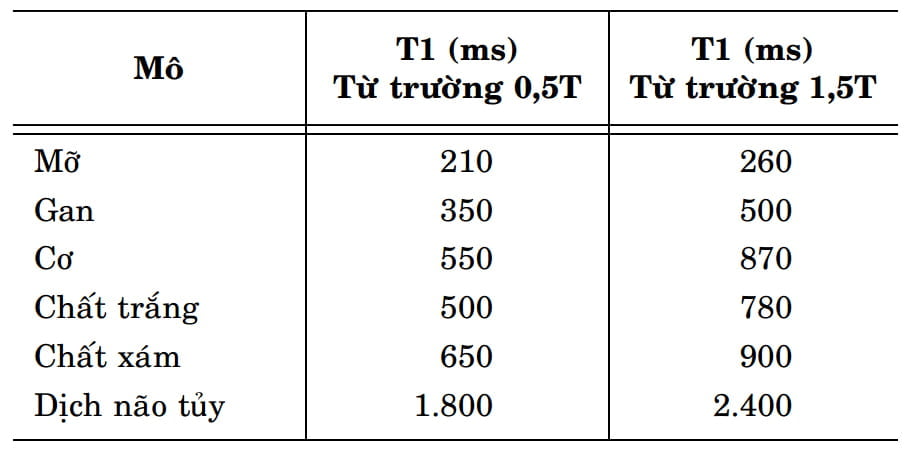
Hình 4: Giá trị T1 tính bằng đơn vị ms (mili-giây) của các mô khác nhau được đo trong hai từ trường với cường độ là 0,5T và 1,5T. Bảng này cũng cho thấy rằng khi cường độ từ trường tăng, thời gian T1 cũng tăng theo tuy mức độ tăng không đồng đều.
Theo chiều hướng đó, các loại dịch tiết trong cơ thể như dịch khớp và mủ có thời gian T1 dài hơn so với T1 của các mô mềm. Tuy nhiên nếu so với các dịch thấm như dịch não tủy và nước tiểu chẳng hạn, thời gian T1 của loại dịch thấm thường dài hơn đáng kể so với các loại dịch tiết vì chúng chứa rất ít các phân tử lớn.
Cũng cần ghi nhận rằng nước tự do trong máu cũng khá lớn nên T1 của máu không ngắn hơn nhiều so với T1 của dịch não tủy. Tuy nhiên do ảnh hưởng mạnh của dòng chảy trong máu nên thời gian T1 của máu không còn được dùng làm một tham số đặc trưng như trong các cấu trúc tĩnh. Các phần tiếp theo sẽ bàn luận chi tiết về hiệu ứng dòng chảy (flow effect) và những ứng dụng của nó trong lĩnh vực cộng hưởng từ tim mạch.
Ảnh hưởng của cường độ từ trường
Như chúng ta đã thấy trong Hình 4, khi cường độ từ trường tăng lên, thời gian T1 của các mô có xu hướng tăng theo mặc dù không đồng đều. Mô mỡ tăng rất ít, chỉ từ 210 ms ở từ trường 0,5T thành 260 ms ở từ trường 1,5T (tỷ lệ 260/210 ≈ 1,2). Các mô mềm đều tăng đáng kể, chẳng hạn tỷ lệ tăng của mô cơ là 870/550 ≈ 1,6. Dịch não tủy, do có lượng nước cao nên có thời gian T1 thay đổi ít hơn (tỷ lệ 2400/1800 ≈ 1,3).
Khác biệt thời gian T1 giữa các mô
Trong các ứng dụng lâm sàng, khác biệt thời gian T1 giữa các mô thường được tận dụng để tạo hình cộng hưởng từ, cho phép chúng ta phân định rõ ranh giới giữa chúng. Nghĩa là trên hình cộng hưởng từ, các mô có thời gian T1 khác nhau càng lớn thì mức độ trắng đen giữa chúng càng rõ mà theo cách nói trong nghề “hình ảnh”, chúng có độ tương phản (contrast) cao. Hình cộng hưởng từ sử dụng mức độ khác biệt T1 của các mô để tạo độ tương phản được gọi là hình trọng T1 (T1-weighted image). Hình 5 là một hình trọng T1 cho thấy rõ cấu trúc các mô. Độ tương phản và hình trọng T1 sẽ được thảo luận trong trong các phần tiếp theo.
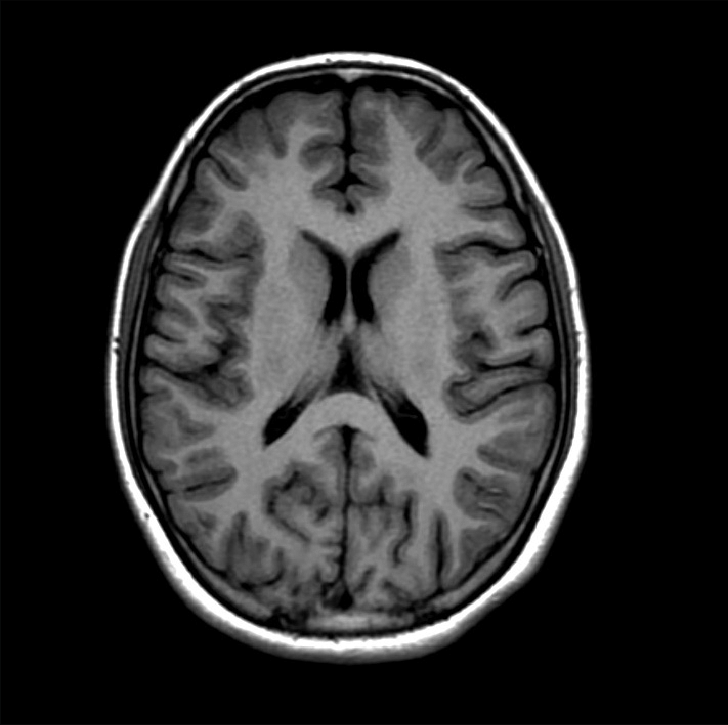
Hình 5: Một hình trọng T1 chụp cắt ngang qua đầu cho thấy rõ các cấu trúc của các mô (chất trắng, chất xám, mô mỡ…).
4. THỜI GIAN HỒI GIÃN NGANG T2
Bây giờ chúng ta quay lại tình huống đã được mô tả ngay từ đầu Phần 2.3 mà ở đó, ngay sau khi tắt xung kích thích, độ từ hóa ngang đang quay quanh trục z với tần số quay bằng tần số quay của các proton. Do hiện tượng cảm ứng điện từ, độ từ hóa ngang này tạo ra một tín hiệu cộng hưởng từ có thể đo được bằng các thiết bị ghi nhận thích hợp.
Bắt đầu từ lúc này, ngoài hiện tượng hồi giãn dọc xảy ra dưới tác dụng của từ trường ngoài B0 đã được thảo luận trong phần trước, hai hiện tượng nhân quả đáng chú ý nữa xảy ra song song với hiện tượng hồi giãn dọc là:
- Quá trình hồi giãn ngang, trong đó độ từ hóa ngang giảm dần độ lớn rồi mất hẳn.
- Do độ từ hóa ngang giảm dần, tín hiệu cộng hưởng từ cũng giảm dần rồi mất hẳn. Đây được gọi là hiện tượng suy giảm cảm ứng tự do FID (Hình 6).
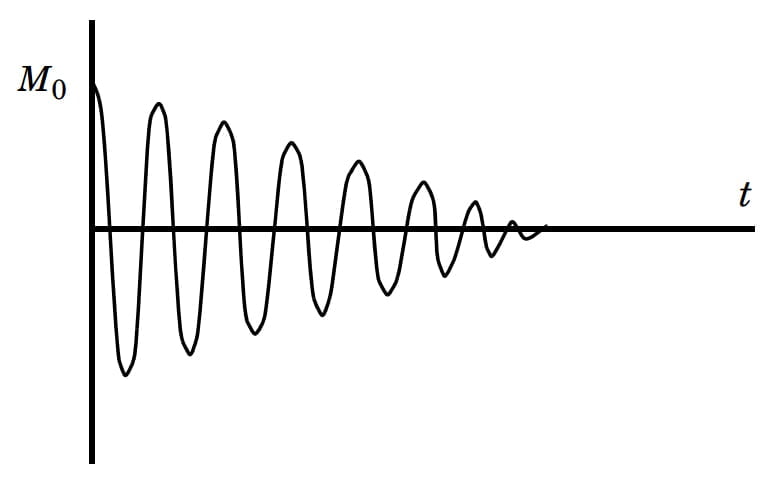
Hình 6: Hiện tượng suy giảm cảm ứng tự do FID: theo thời gian, tín hiệu cộng hưởng từ lúc đầu có độ lớn bằng M0 giảm dần rồi mất hẳn.
Trong khi cơ chế của quá trình hồi giãn dọc là sự tương tác giữa proton với môi trường (tương tác spin-lattice) xảy ra dưới tác dụng của từ trường ngoài B0, cơ chế của quá trình hồi giãn ngang lại do tương tác giữa các proton với nhau (tương tác spin-spin). Lúc đầu các proton quay cùng pha, tạo ra một “sức mạnh tổng hợp” tối đa. Theo thời gian, các proton quay gần nhau có thể “đụng nhau” (tác động từ tính qua lại) khiến cho chúng dần lệch pha, làm giảm bớt đi sức mạnh tổng hợp của chúng. Nghĩa là vectơ từ hóa ngang lúc này nhỏ hơn so với lúc đầu.
Đường cong của quá trình hồi giãn ngang
Tương tự như quá trình hồi giãn dọc, người ta cũng mô tả quá trình hồi giãn ngang bằng một đường cong (Hình 7). Đường cong này cho thấy mức độ hồi giãn ngang, tức là sự suy giảm của vectơ từ hóa ngang, lúc đầu xảy ra khá nhanh rồi chậm dần. Ở đây, thời gian hồi giãn ngang hay thời gian T2 được tính là khoảng thời gian để vectơ từ hóa ngang giảm đi khoảng 63% độ lớn của mình hay nói cách khác, nó chỉ còn khoảng 37% độ lớn.
Chúng ta cần nhớ rằng tín hiệu cộng hưởng từ được tạo ra do độ từ hóa ngang, vốn là một từ trường, quay quanh trục z. Do vậy, suy giảm vectơ từ hóa ngang sẽ gây ra hiện tượng suy giảm cảm ứng tự do FID và thời gian T2 cũng chính là thời gian suy giảm tín hiệu cộng hưởng từ. Mặt khác, khi vectơ từ hóa dọc được khôi phục hoàn toàn thì vectơ từ hóa ngang chắc chắn không còn tồn tại nữa. Vì thế thời gian T1 có thể được xem như giá trị lớn nhất của T2 (trong trường hợp nước tinh khiết). Thực tế, T2 thường nhỏ hơn nhiều so với T1.
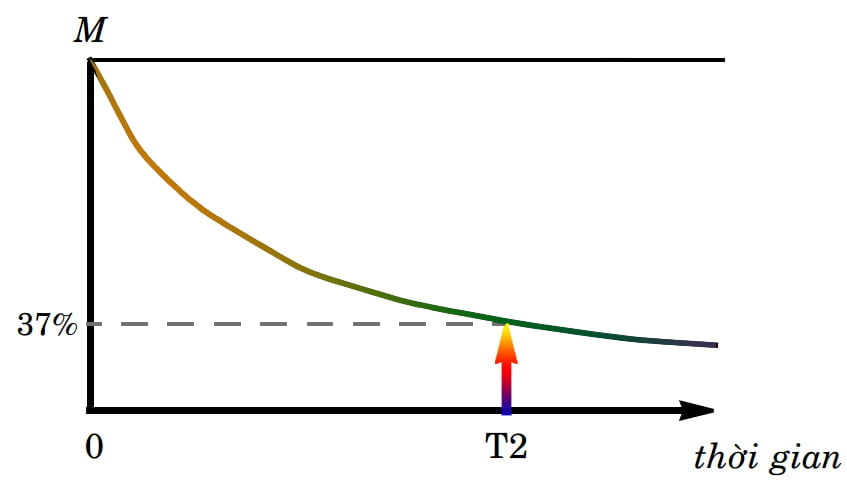
Hình 7: Đường cong mô tả thời gian hồi giãn ngang cho thấy mối liên hệ giữa độ lớn của vectơ từ hóa ngang với thời gian. T2 là thời gian vectơ từ hóa ngang chỉ còn khoảng 37% độ lớn so với độ lớn ban đầu M của nó. Nghĩa là hết một thời gian T2, nó đã mất khoảng 63% độ lớn của mình.
Thời gian T2 của một số mô điển hình
So với thời gian T1, thời gian T2 của các mô cơ thể thường ngắn hơn nhiều. Bảng được đưa ra trong Hình 8 minh họa cho chúng ta thấy điều này. Vì thời gian T2 biểu thị tốc độ suy giảm tín hiệu, giá trị T2 trong bảng đó giúp chúng ta so sánh độ suy giảm tín hiệu của các mô khác nhau. Với các mô có thời gian T2 ngắn, tín hiệu sẽ suy giảm nhanh và ngược lại với các mô có thời gian T2 dài, tín hiệu sẽ tồn tại lâu hơn. Do vậy theo bảng này, gan là mô có tín hiệu suy giảm nhanh nhất; dịch não tủy là mô có tín hiệu suy giảm lâu nhất.
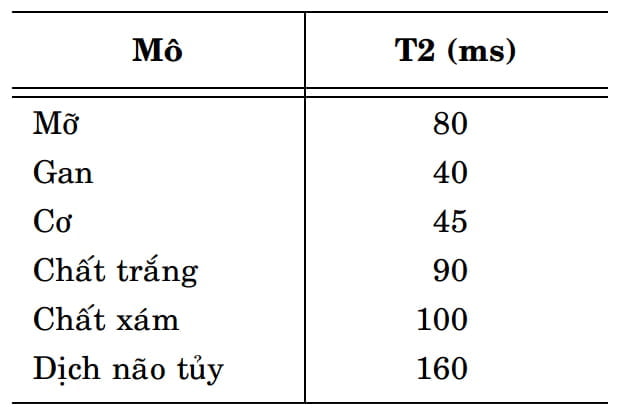
Hình 8: Giá trị T2 tính theo đơn vị ms (mili-giây) của các mô khác nhau được đo trong một nghiên cứu giống như bảng của Hình 4. Nếu so sánh với giá trị T1 của bảng được cho trong Hình 4, giá trị T2 của các mô tương ứng nhỏ hơn nhiều, chỉ nằm trong khoảng từ vài chục đến vài trăm mili-giây.
Chúng ta cần nhớ rằng các mô trong cơ thể vốn phức tạp do chứa nhiều loại tế bào và các chất chuyển hóa khác nhau. Ở mức vi thể, chúng đều có những từ trường riêng rất nhỏ tác động qua lại, tạo ra một từ trường cục bộ không đồng nhất. Đây là một nguyên nhân làm cho các proton bị lệch pha ngoài nguyên nhân tương tác trực tiếp giữa các proton đã nêu.
Cụ thể, thời gian T2, tương tự như T1, cũng phụ thuộc vào tỷ lệ nước tự do/ nước tù trong các mô. Tuy nhiên sự phụ thuộc này lại có nguồn gốc từ sự không đồng nhất của từ trường cục bộ vừa nêu. Nước tự do có các phân tử chuyển động nhanh nên các proton của chúng ít bị tác động bởi tình trạng không đồng nhất của từ trường cục bộ xung quanh. Ngược lại nước tù với các phân tử bị giữ chặt bởi các phân tử lớn khiến chúng dễ bị tác động của từ trường cục bộ này, làm ngắn thời gian T2 của những mô có lượng nước tù nhiều. Vỏ xương là một thí dụ rõ nhất. Tín hiệu của vỏ xương hầu như không có do vỏ xương hầu như không có nước tự do. Nước có mặt trong vỏ xương đều là nước tù và bị giữ quá chặt khiến cho thời gian T2 của vỏ xương cực ngắn và hầu như trong mọi chuỗi xung đều không ghi nhận được tín hiệu cộng hưởng từ.
Như vậy, dù căn nguyên là gì đi chăng nữa, cả thời gian T1 và T2 đều phụ thuộc vào tỷ lệ giữa nước tự do và nước tù của các mô. Nhận xét vừa nêu cho phép chúng ta khẳng định rằng trong thực tế, các mô có thời gian T1 dài thường cũng có thời gian T2 dài và ngược lại.
Thời gian hồi giãn ngang T2*
Theo như đã mô tả trong phần trước, quá trình hồi giãn ngang xảy ra do sự tương tác giữa các proton, làm cho vectơ từ hóa ngang suy giảm dần trong thời gian T2. Tuy nhiên trong môi trường cơ thể, tín hiệu cộng hưởng từ thường mất khá nhanh chứ không tồn tại và kéo dài trong suốt thời gian T2, nghĩa là thời gian hồi giãn ngang thực tế này ngắn hơn so với thời gian hồi giãn ngang T2. Để phân biệt rõ ràng thời gian T2 thực tế với thời gian T2, người ta dùng khái niệm thời gian hồi giãn ngang T2*.
Lý do giải thích sự suy giảm nhanh chóng tín hiệu cộng hưởng từ này được quy cho các nguyên nhân đã dẫn đến tình trạng không đồng nhất của từ trường cục bộ và độ xê dịch hóa học vốn có giữa các proton của mỡ và của nước. Tính không đồng nhất của từ trường cục bộ, xét ở một góc độ nào đó, là một nguyên nhân vốn có vì như đã giải thích trong phần trước, các mô cơ thể có rất nhiều chất với những từ trường khác nhau. Ngoài ra, các chất thuận từ được đưa từ ngoài vào (chẳng hạn như các thuốc tương phản từ) hay được tạo ra trong một quá trình bệnh lý cũng là một nguyên nhân gây ra tính không đồng nhất của từ trường cục bộ.
Cũng cần nhấn mạnh rằng thuật ngữ T2 được dùng với ý nghĩa thông thường là thời gian hồi giãn ngang biểu thị một đặc trưng vốn có của mỗi mô cơ thể. Trong trường hợp cần nói đến ảnh hưởng của những nguyên nhân khác, chẳng hạn ảnh hưởng của tình trạng không đồng nhất của từ trường cục bộ hoặc do độ xê dịch hóa học, người ta dùng T2*. Điều này có nghĩa là T2* được dùng thay cho T2 trong những trường hợp đặc biệt với những mục đích đặc biệt.
Ảnh hưởng của độ xê dịch hóa học
Như chúng ta đã biết từ Phần 1, độ xê dịch hóa học là sự khác biệt về tần số quay giữa các proton ở những phân tử khác nhau. Với những kỹ thuật cộng hưởng từ hiện có, proton của nước và của mỡ là hai nguồn quan trọng nhất tạo ra tín hiệu cộng hưởng từ.
Trước tiên, bản thân mô mỡ chứa nhiều loại phân tử với những độ xê dịch hóa học khác nhau, mặc dù nguồn tín hiệu quan trọng nhất của mô mỡ là các proton CH2 bão hòa trong các phân tử triglyceride.
Thứ hai, proton của nước có tần số quay hơi nhanh hơn so với proton của CH2. Vì thế mặc dù lúc đầu khi vừa tắt xung kích thích, các proton của nước và của CH2 đều quay cùng pha nhưng theo thời gian, do ảnh hưởng của độ xê dịch hóa học, chúng dần lệch pha nhau cho đến thời điểm pha của chúng chênh lệch nhau 180o (nghịch pha). Lúc này tín hiệu hoàn toàn bị biến mất. Thế nhưng vì proton của nước vẫn quay nhanh hơn proton của CH2 nên mất một khoảng thời gian bằng như thế nữa, proton của nước và của CH2 sẽ có pha chênh nhau 360o. Tại thời điểm này, chúng lại cùng pha và tín hiệu lúc này mạnh trở lại. Đây là cơ sở của kỹ thuật Dixon thực hiện xóa tín hiệu của mỡ sẽ được phân tích trong các phần tiếp theo.
Như vậy, độ xê dịch hóa học không hoàn toàn làm mất hẳn tín hiệu cộng hưởng từ: nó chỉ làm thay đổi tín hiệu cộng hưởng từ theo một quy luật nhất định. Quy luật này sẽ được sử dụng trong nhiều trường hợp để gợi ý hoặc khẳng định chẩn đoán.
Ảnh hưởng của các chất thuận từ
Chúng ta đã biết rằng chất thuận từ là nhóm chất có khả năng bị từ hóa dưới tác động của một từ trường. Khi đó chúng trở thành một nam châm yếu và từ trường của chúng hợp sức với từ trường hiện có, làm cho từ trường chung mạnh hơn, mặc dù so với tác động của nhóm chất sắt từ, sức mạnh bổ sung này khá nhỏ.
Tuy nhiên ở mức vi thể, chất thuận từ có thể làm mất tính đồng nhất của từ trường cục bộ, một nguyên nhân quan trọng làm suy giảm nhanh tín hiệu cộng hưởng từ, khiến cho thời gian T2* vốn đã ngắn lại càng ngắn hơn. Tuy nhiên ảnh hưởng này phụ thuộc vào mức độ tiếp xúc giữa chất thuận từ với các phân tử nước. Một chất thuận từ có thể có rất ít tác động đối với mô nếu nó bao khuất không nằm sát với các phân tử nước.
Tùy theo hàm lượng và mức độ thuận từ, một chất thuận từ khi có tác động của một từ trường ngoài có thể làm tăng cường độ của từ trường cục bộ. Những chất thuận từ đó đôi khi được gọi bằng thuật ngữ chất nhạy từ (superparamagnetic) và được định nghĩa là những chất có khả năng từ hóa giữa mức chất sắt từ và chất thuận từ. Chẳng hạn các loại thuốc tương phản từ khi được đưa vào bằng đường tĩnh mạch đều nằm trong nhóm chất nhạy từ. Với một nồng độ cao và tiếp xúc đều khắp với các phân tử nước, các loại thuốc này làm tăng cường độ từ trường cục bộ, kết quả là cả thời gian T1 và T2 đều ngắn đi.
Một số quá trình bệnh lý có thể sản sinh ra các chất thuận từ. Thí dụ như trong quá trình phân hủy cục máu đông, một sản phẩm phân hủy hemoglobin của hồng cầu là chất methemoglobin, vốn là một chất thuận từ, có thể làm thay đổi tín hiệu của vùng bị xuất huyết. Trong giai đoạn bán cấp sớm, chất methemoglobin được tạo ra nhưng hồng cầu chưa bị vỡ nên ảnh hưởng của nó không rõ ràng. Tuy nhiên ở giai đoạn bán cấp muộn khi hồng cầu đã vỡ, methemoglobin làm thay đổi rõ rệt tín hiệu cộng hưởng từ.
Khác biệt thời gian T2 giữa các mô
Tương tự như T1, khác biệt thời gian T2 giữa các mô cũng được tận dụng để tạo sự tương phản giữa các mô trên hình cộng hưởng từ. Một hình cộng hưởng từ có sử dụng sự khác biệt thời gian T2 giữa các mô được gọi là hình trọng T2 (T2-weighted image): khác biệt T2 của hai mô càng lớn, mức độ trắng đen giữa chúng trên ảnh trọng T2 càng rõ. Hình 9 trình bày một hình trọng T2 của não cho thấy cấu trúc các mô không rõ ràng lắm. Tuy nhiên tín hiệu của dịch não tủy rất mạnh, thể hiện bằng hai vùng trắng tương ứng với hai não thất bên và các đường trắng len giữa các hồi não. Độ tương phản và hình trọng T2 cũng sẽ được thảo luận trong các phần tiếp theo.
5. NHỮNG ĐIỂM CẦN GHI NHỚ
Đến đây chúng ta đã nghiên cứu xong những đặc điểm quan trọng của hiện tượng cộng hưởng từ trên các mô cơ thể. Chúng là cơ sở để trong những phần sau, với nhiều kỹ thuật cực kỳ độc đáo, chúng ta có thể “nhìn thấu” vào trong cơ thể sống bằng cách tạo ra những hình ảnh giống như thể chúng ta đang cắt cơ thể sống ra thành từng lớp để biết được chúng đang “sống” như thế nào. Những khái niệm quan trọng và đáng chú ý trong phần này được tóm tắt như dưới đây:
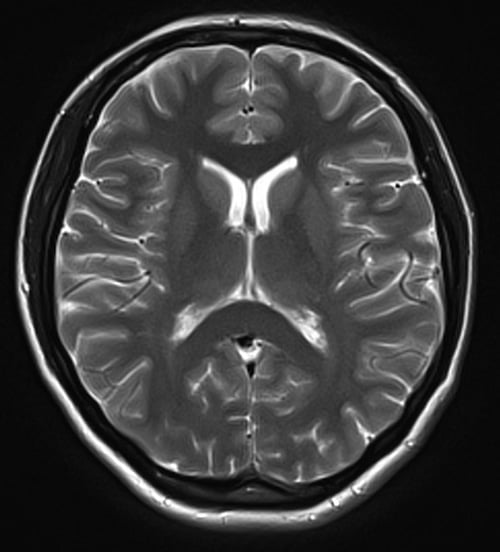
Hình 9: Một hình trọng T2 chụp cắt ngang qua đầu ngang mức não thất bên cho thấy rõ hình dạng của hai não thất bên nhờ chứa nhiều dịch não tủy. Phân biệt giữa chất xám và chất trắng không được rõ ràng.
- Hình ảnh cộng hưởng từ được tạo thành từ hai nguồn tín hiệu quan trọng của cơ thể là nước và mỡ. Các proton của nước có tần số quay nhanh hơn chút ít so với tần số quay của các proton trong mỡ. Khác biệt này thường được gọi là độ xê dịch hóa học.
- Nước là thành phần chủ yếu có mặt khắp nơi trong cơ thể. Về mặt tín hiệu cộng hưởng từ, chúng ta chỉ phân biệt hai loại nước trong cơ thể: nước tù và nước tự do.
- Nước tù là phần nước nằm gần các phân tử lớn. Những phân tử nước ở tình trạng này bị các đại phân tử hút chặt nên chúng chuyển động rất ít, do vậy chúng dễ bị tác động của từ trường.
- Ngược lại, nước tự do chuyển động rất nhanh vì chúng ở xa các đại phân tử. Nhờ tốc độ chuyển động như thế, nước tự do bị tác động của từ trường chậm hơn nước tù.
- Quá trình hồi giãn dọc xảy ra do sự tương tác giữa proton với môi trường dưới tác dụng của từ trường ngoài. Thời gian hồi giãn dọc còn được gọi là T1.
- Quá trình hồi giãn ngang và hiện tượng suy giảm cảm ứng tự do FID là hai hiện tượng nhân quả xảy ra song song với quá trình hồi giãn dọc. Thời gian hồi giãn ngang còn được gọi là T2 và thường ngắn hơn nhiều so với T1.
- Tín hiệu cộng hưởng từ của mô thường suy giảm nhanh hơn so với thời gian T2 của nó. Nguyên nhân của nó được quy cho tình trạng không đồng nhất của từ trường cục bộ và độ xê dịch hóa học của các mô. Khi đó, T2 được gọi là T2*.
- Tỷ lệ giữa nước tự do và nước tù là một dấu chỉ quan trọng cho tính chất cộng hưởng từ của mô. Các mô mềm có lượng nước tù nhiều hơn nên thời gian T1 và T2 thường ngắn. Các loại dịch tiết có lượng nước tự do nhiều hơn nên T1 và T2 dài hơn. Tuy nhiên các loại dịch thấm luôn có T1 và T2 dài nhất vì có lượng nước tự do nhiều nhất.
- Khác biệt thời gian hồi giãn giữa các mô có thể được dùng để phân định cấu trúc của chúng trên hình cộng hưởng từ. Một hình dùng T1 để phân định cấu trúc của các mô được gọi là hình trọng T1. Tương tự, một hình dùng T2 để phân định cấu trúc của các mô được gọi là hình trọng T2.